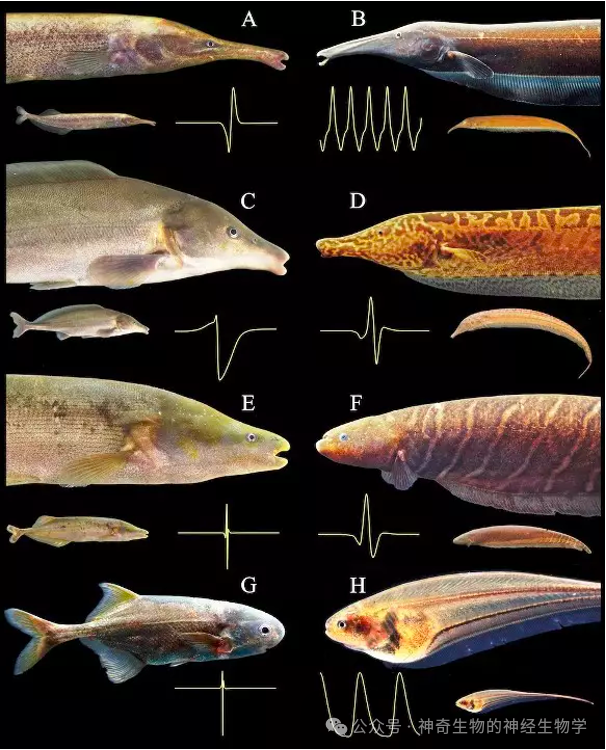
说到会放电的鱼,我们通常会想到能都把牛都电晕的电鳗和电鳐。这些可怕的鱼类通过释放几百伏高压电来捕食和防御,可以说在自然界无人能敌。而我们介绍的则被称为弱电鱼,他们放出的电压通常1V左右,不具有杀伤力,而是用来感知环境和用来与同类交流。
弱电鱼有两大类:
(1)魔鬼刀鱼,来自南美,产生正弦波型电信号;
(2)象鼻鱼,来自非洲,产生脉冲型电信号。
从外形上看,两大类弱电鱼的不同,尾巴有没有分叉,鳍是否从头到尾延伸。本质上是游泳方式的不同。

但不完全如此,非洲象鼻鱼中有些类群的放电是正弦波型,这些鱼就长得和刀鱼很像(被称为反飞刀鱼,波动鳍在背部):这就体现了体型与信号的紧密关系。脉冲型电鱼和普通鱼类一样摆动尾巴游泳;正弦波型电鱼通过鳍波动游泳,是为了使得身体可以保持不动。南美飞刀鱼,也有不少脉冲型。

为什么研究电鱼?
首先,因为可以使用简单的刺激:在那个还没有电脑可以生产复杂刺激的年代(1970s),人工生成简单正弦波就足以骗过电鱼。
其次,放电作为一个晚近演化出的新功能,很可能有一个显眼的,与众不同的神经结构。
对于电鱼的研究有几个先驱性的重要工作,主要由Scripps研究所的神经生物学家Ted Bullock开启,由他的学生Walter Heiligenberg完成:
(1)JAR(Jamming avoidance reflect):用于定位时,正弦波型会进行电波调频错峰。在脊椎动物中,第一个完整解释从感觉输入到**输出完整神经机制的系统。
(2)交流信号中的时间编码:脉冲型电鱼;神经系统(包括听觉、视觉等)中时间信息处理通用算法。
(3)感知副本(efference copy,又称corollary discharge):脉冲型电鱼交流时,感觉系统主动过滤掉自己发出电信号;在神经算法层面上第一次完整地解释电鱼在交流的过程中的感知副本是如何生成的。
电波的产生与感知
很多动物可以感受环境的电场,称为电感受(eletroreception)。
这种能力在自然界中并不罕见。七鳃鳗,鲨鱼,硬骨鱼,蝾螈,甚至是哺乳动物里的鸭嘴兽,大部分的水生动物都具备这种能力。
因为水可以导电,而每当动物**的时候,肌肉收缩会伴随着肌肉电的产生,通过感受其他动物靠近时发出的肌肉电,水生动物可以发现隐蔽在黑暗中的猎物或者天敌,称为被动电场定位 (passive electrolocation)。正因为如此有用,生物进化的历程中,经常伴随着到陆地生活,失去感受电场的能力(因为空气不易导电);回到水里生活,又重新进化出感受电场的能力,如此循环。
像这样被动地感受别的动物发出的肌肉电有一个缺点,就是如果猎物或者天敌埋伏不动,不产生肌肉电,就无法探测到它们。
于是机智的非洲电鱼和南美电鱼在地球的两端不约而同地进化出了一种技能,主动的产生一个电场。肌肉产生肌肉电本身是为了刺激其收缩,电鱼的尾部都一些肌肉发生特殊变化,只产生肌肉电而不收缩,变成了单纯的发电器官(electric organ)。
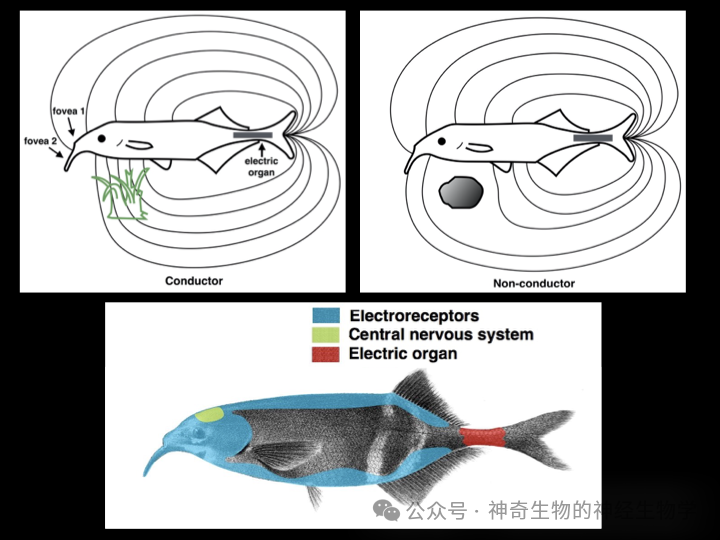
而分布在电鱼身体和头部各处的电场感受细胞则可以检测尾部发出的电场变化。于是,它就可以像一个活体金属探测器一样搜索水底,当它的身边某处有可以导电性生物的时候,那里附近的电场感受细胞会感觉到更大的电流。相反,如果那里是一块不能导电的石头,那里附近的电场感受细胞会感觉到更弱的电流。通过感受自己产生的电场在环境里被变形的状态,电鱼就可以在漆黑的夜里,或者浑浊的水域“看”到水底的障碍物和隐匿的猎物。这种能力称为主动电场定位 (active electrolocation)。
正弦波型电鱼之所以需要游泳的时候保持身体不动,就是因为它持续放电,通过电场感知环境。一旦摆动身体游泳,产生的电场形状就发生变化,干扰电场感知。而脉冲型电鱼因为是瞬时发出的是离散脉冲,所以摆动身体游泳影响不大。
剑桥大学的Hans Lissmann在1950年代通过训练动物区分不同电导率的物体,不同盐度的水瓶,和食物奖赏或者惩罚相关联,首次证明鱼类具有感受电场的‘第六感’。
鱼类具有感受电场的电感受器(electroreceptor)有不同类型:
(1)壶腹型(Ampullary type),是比较原始的,被动感知低频的微弱的电信号(DC signal):因为很长的导管,使得高频信号被过滤掉。软骨鱼有遍布头部的管线系统。劳伦氏壶腹(ampullae of Lorenzini)是意大利医生Stefano Lorenzini在1678年发现。
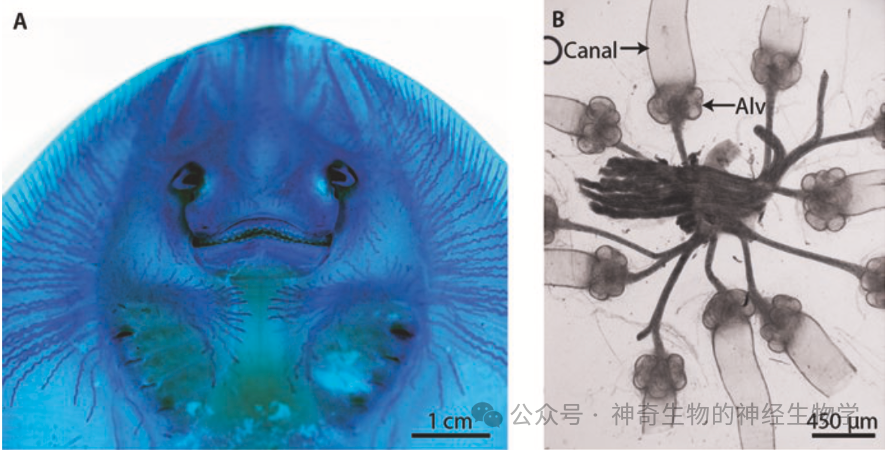
(2)结节型(Tuberous type):弱电鱼所特有,新近演化的,适用于高频的比较强的(主动放电)电信号(AC signal)
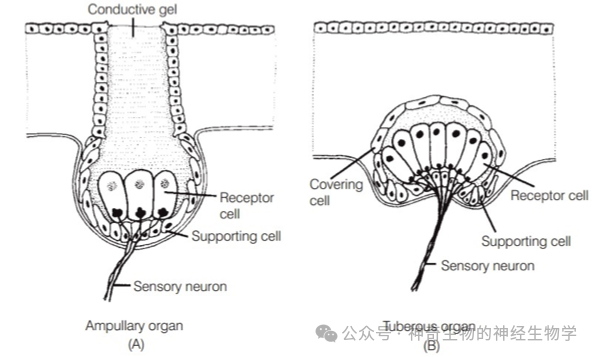
它们都演化自侧线系统的机械感受器。和陆生动物的听觉器官是同源的。
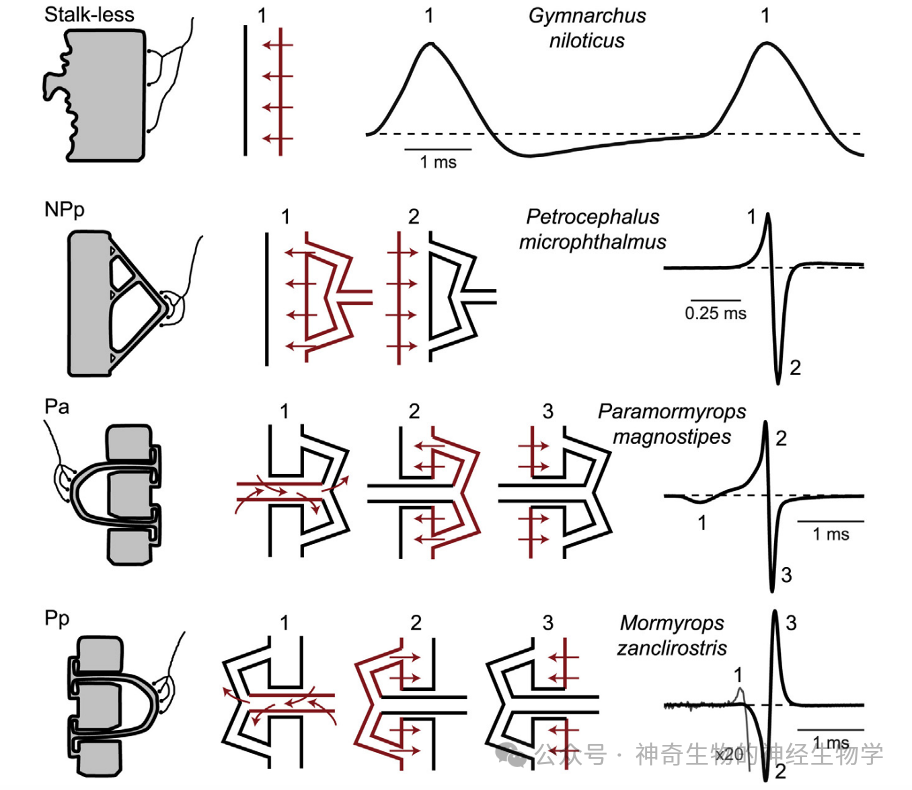
能够主动放电的电鱼具有发电器官(electric organ)产生EOD (electric organ discharge): 它是特化的肌肉细胞,每个肌肉细胞产生几十mV电压,规则排列起来(串联)叠加超过1V的放电。不同的肌肉与神经的布线方式使得不同物种的电鱼产生的EOD有不同的波形。
电信号处理中枢:全身表面的电感受器形成“电成像”(electric image:seeing with the body surface),这些信息通过传入神经汇入位于后脑的电感侧线核团ELL(electrosensory lateral line lobe )。ELL具有多个身体地图(somatotopic map):内侧一个来自壶腹型电感受器(Ampullary);外侧3个来自结节型电感受器(Tuberous);和我们之前见到的身体地图一样,保持拓扑关系,当表征不平均:头部占据更大区域(over-represented)。
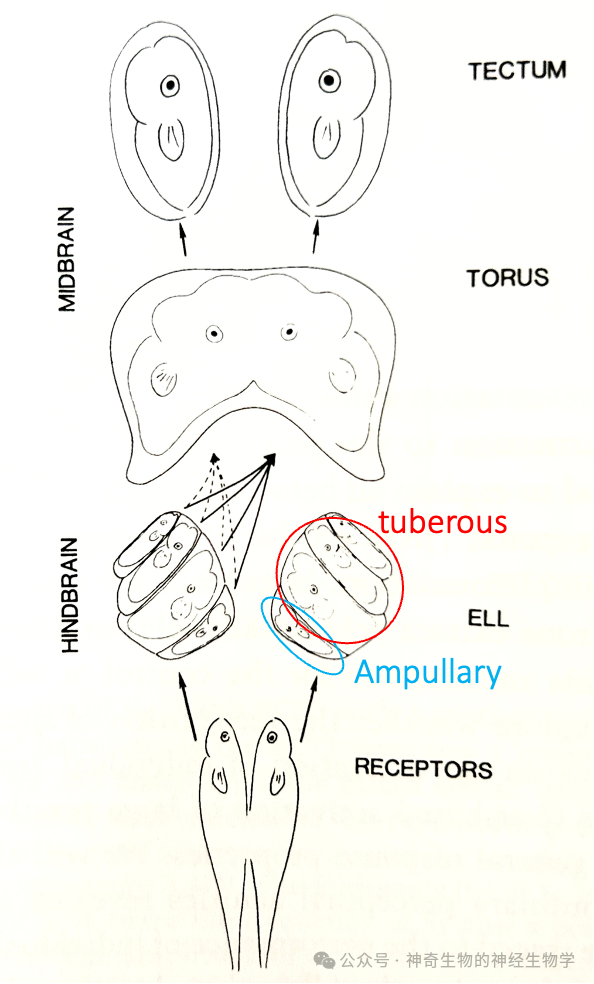
ELL的输出投射到中脑的torus semicircularis (对应四足动物脑的下丘inferior colliculus)汇合成一个统一的身体地图;再投射到optic tectum。这条通路和猫头鹰用于声音定位的神经通路很像,因为电感受器演化来自侧线系统,和陆生动物的听觉是同源的。
电波调频避让反应JAR
一只电鱼可以愉快地收听自己放出的电波在环境里的“回响”,但两只电鱼同时在放电的时候问题就来了。
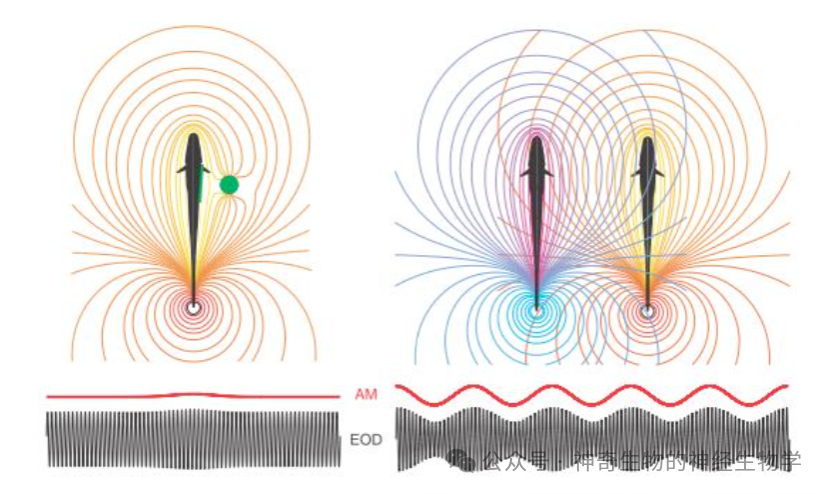
每只电鱼通常以接近但不完全相同的频率放出电波,这两个电波就会发生干涉。干涉使得每一只鱼感受到的不再只是自己发出的电波,而是一个振幅呈周期性变化的叠加电波(调幅的频率等于两个波源频率之差),而这个振幅变化的幅度远大于它们用来探测到身旁导电物体所以根据的幅度变化,于是这个活体金属探测器就受干扰失灵了。
这就像是不同广播电台会有自己固定的频率,如果两个电台频率太接近就会互相串台。而电鱼应对这种干扰的策略是在频道上相互礼让:原本放电频率高的再稍微调高一点,原本放电频率低的再稍微调低一点。这个行为叫做JAR(Jamming avoidance response)
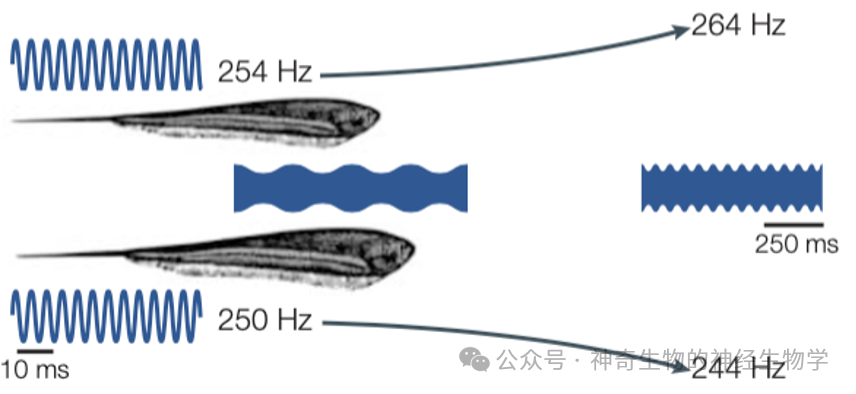
这个相互礼让的方法听起来容易,但做起来并不容易。关键在于,电鱼需要判断另外一只电鱼的放电频率,究竟是比自己高,还是比自己低。如果这个判断错了,就像迎面开来的两辆车,一辆往左让,一辆往右让,结果就撞在一起了。
计算层面 (Computation):判断对方放电频率是否比自己高
为了实现放电频率避让,一个简单有效的方法是与自己的放电频率做比较。但是,进化不一定总能找到最有效的办法。同时,内在的世界模型是很难实现的。
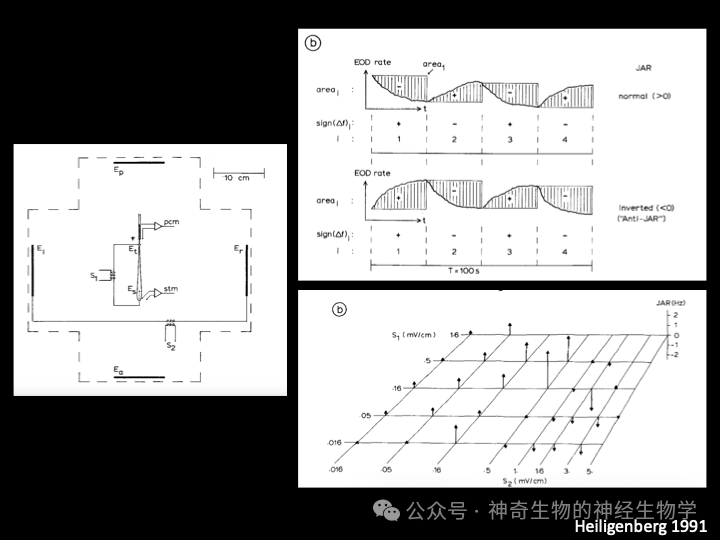
行为实验证明(the experimental opening of a loop),电鱼在进行JAR时,不知道自己的放电频率:首先使用箭毒阻断神经对肌肉的控制(NMJ nAChR),在鱼头尾放置电极(S1)模拟其自身放电,同时记录pacemaker(medulla oblongata决定发电器官放电频率的起搏器神经细胞)的放电频率,远处电极(S2)模拟另一只鱼放电。(1)如果以记录到的pacemaker的频率赋予S1模拟其自身放电,改变S2(另一只假鱼)放电频率,可以诱发pacemaker的放电频率产生JAR。(2)把pacemaker的放电频率减去50Hz赋予S1代替其自身放电,JAR则以-50Hz为自身频率,而非pacemaker的频率。(3)如果S1的放电强度低于S2的放电强度,甚至会引起反方向的JAR(anti-JAR)但是,如果仅仅用一对电极模拟两个频率的干扰(回放两波叠加信号),也不能引起JAR。因为这时候全身各处感受到的两波叠加干扰的程度是一样的(identical geometry)。干扰信号必须从一方来,使得动物身体不同侧的干扰比例不同。
所以,电鱼是在不知道自己的频率的情况下,计算两个干扰电波的频率差的符号,而且默认那个增幅更大的电波是自己的。
要从业已叠加在一起的电波中得知对方的频率高低,电鱼采用的方法是看叠加电波相位与振幅的关系。
两波叠加信号的差拍周期 (beat cycle,即波包频率)等于二者频率差在身体不同部分干扰的强度不同,垂直体表最强。因为自身产生的电波的波源比较近,所以振幅会比较大。
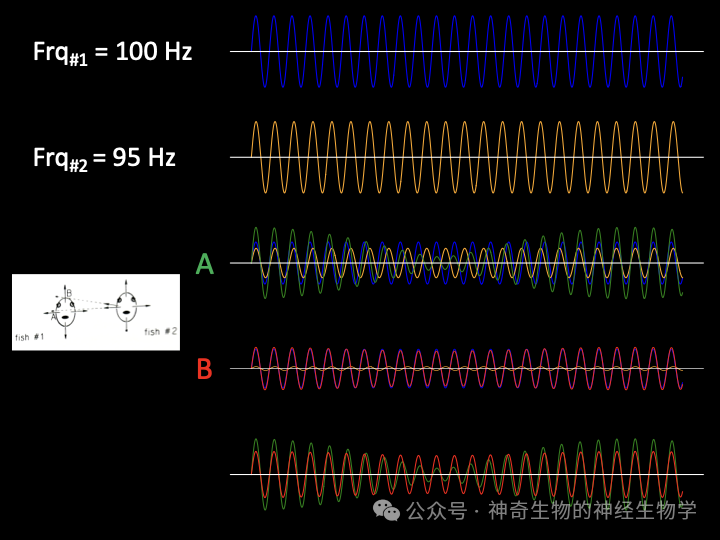
如果对方的频率稍慢(Df<0),叠加起来的结果是:振幅变小的时候,即较慢的频率(对方)主导的时候,相位会推迟(200Hz 一个周期是5ms,相位差小于pi/2<1ms);振幅变大的时候,即较快的频率(己方)主导的时候,相位会推提前。这种情况下,自己的放电频率应该上调。
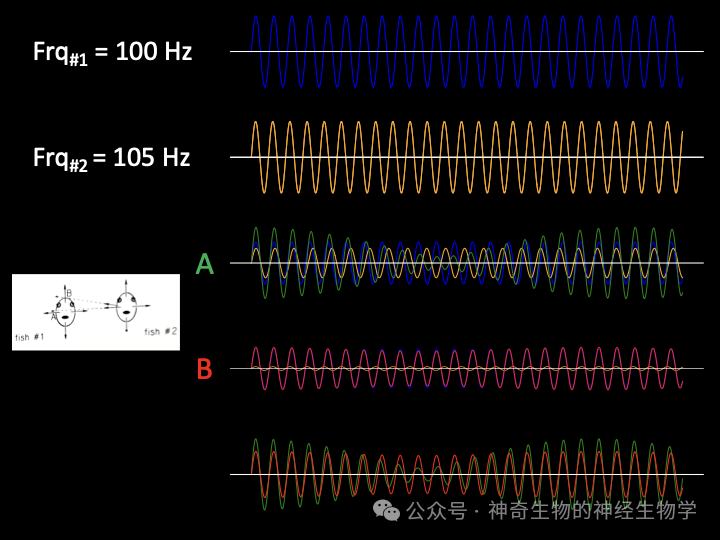
如果对方的频率稍快(Df>0),叠加起来的结果是:振幅变小的时候,即较快的频率(对方)主导的时候,相位会提前;振幅变大的时候,即较慢的频率(己方)主导的时候,相位会推推迟。这种情况下,自己的放电频率应该下调。
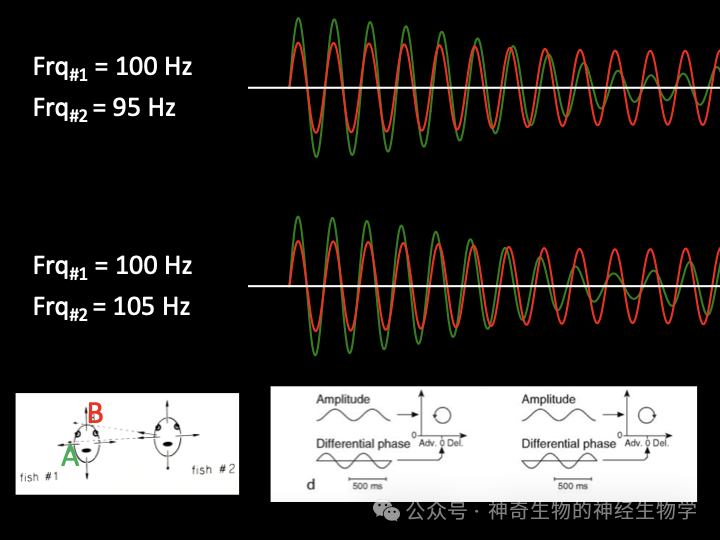
身体部分的独立分析自身的波包振幅和相位-振幅关系,相位-振幅关系决定放电频率改变的方向;波包振幅决定贡献的权重。最终,波包振幅最大的身体区域胜出放电频率改变的方向。
算法与神经实现(Algorithm/nerual implementation)
结节型电感受器(tuberous-type electroreceptor)分为两类:
(1)P-type (probability):编码(波包)振幅强度,调幅(AM)。
(2)T-type (time):编码相位(phase lock to zero crossing),调频(FM)。通过比较身体不同区域的相位,可以得到相位差;
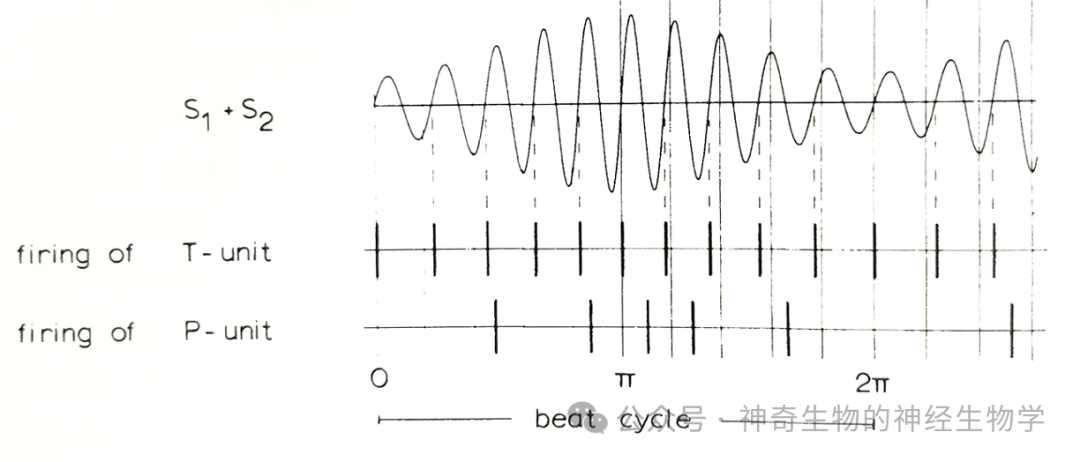
后脑的电感侧线核团ELL(electrosensory lateral line lobe): 具有somatotopic map。有三条神经通路:
(1)T-type传入神经通过电突触投射到球状细胞 (spherical cell),高度同步化多条T-type传入纤维才能激活,增强相位编码的时间精确度。
(2)P-type传入神经激活basilar pyramidal cells,编码振幅增大
(3)P-type传入神经通过granule cells抑制non-basilar pyramidal cells,编码振幅减小
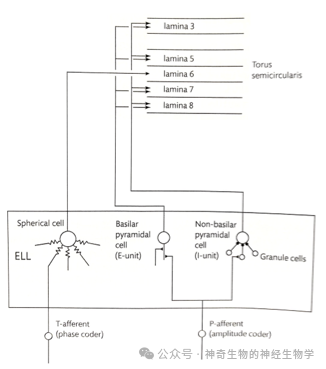
中脑 TS(torus semicircularis,对应四足动物脑的下丘inferior colliculus)的是个多层(lamina)结构的脑区。
其中,第六层(lamina 6)处理时间/相位信息,进行身体各个点的相位关系(pairwise)比较,计算出任意两点的相位关系是提前还是延后,获得一个相位关系矩阵。
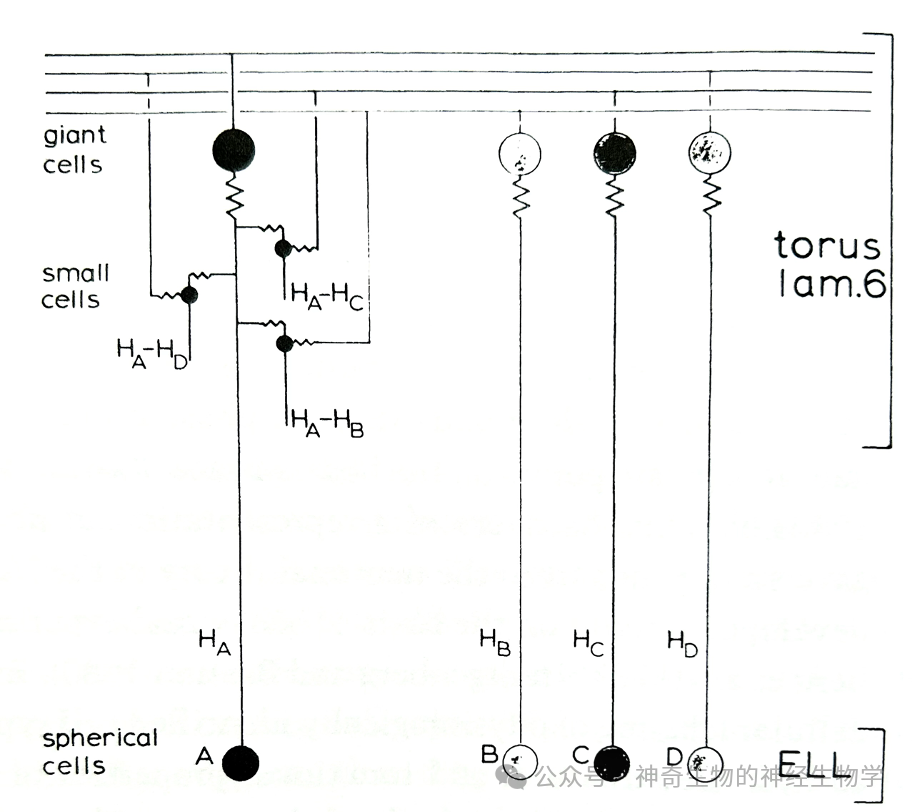
ELL球状细胞 (spherical cell)按拓扑关系(somatotopy)投射到对应区域TS的巨大细胞(giant cell)的胞体(soma)和小细胞(small cell)的树突(dendrite)。
巨大细胞(giant cell)产生平行纤维(parallel fibers)横贯整个TS的第六层,随机连接各处小细胞(small cell)的胞体(soma)。
每个小细胞(small cell)只随机接受一个远处巨大细胞(giant cell)的投射和一个ELL球状细胞 (spherical cell)的投射,后者具有一个按照身体地图分布的身体表面感受野(somatotopic receptive field)。
小细胞(small cell)进行相位比较:细胞所在位置(对应身体表面的一个位点)信号树突上传播有个延迟(delay line),对比该信号和巨大细胞(giant cell)带来的远处(对应身体表面的另外一个位点)的信号。
于是出现在TS第六层出现对相位延迟(delay)敏感的小细胞和对相位提前(advance )敏感的小细胞。
TS其他层(lamina)处理强度/振幅信息。TS跨层的纵向连接是振幅和相位信息汇合。在第八层(lamina 8)通过组合与门实现四类细胞:(1)振幅变小,相位提前;(2)振幅变大,相位提前;(3)振幅变小,相位推迟;(4)振幅变大,相位推迟。
其中,(1)和(4)持续激活,说明对方的频率比我快。(2)和(3)持续激活,说明对方的频率比我慢。
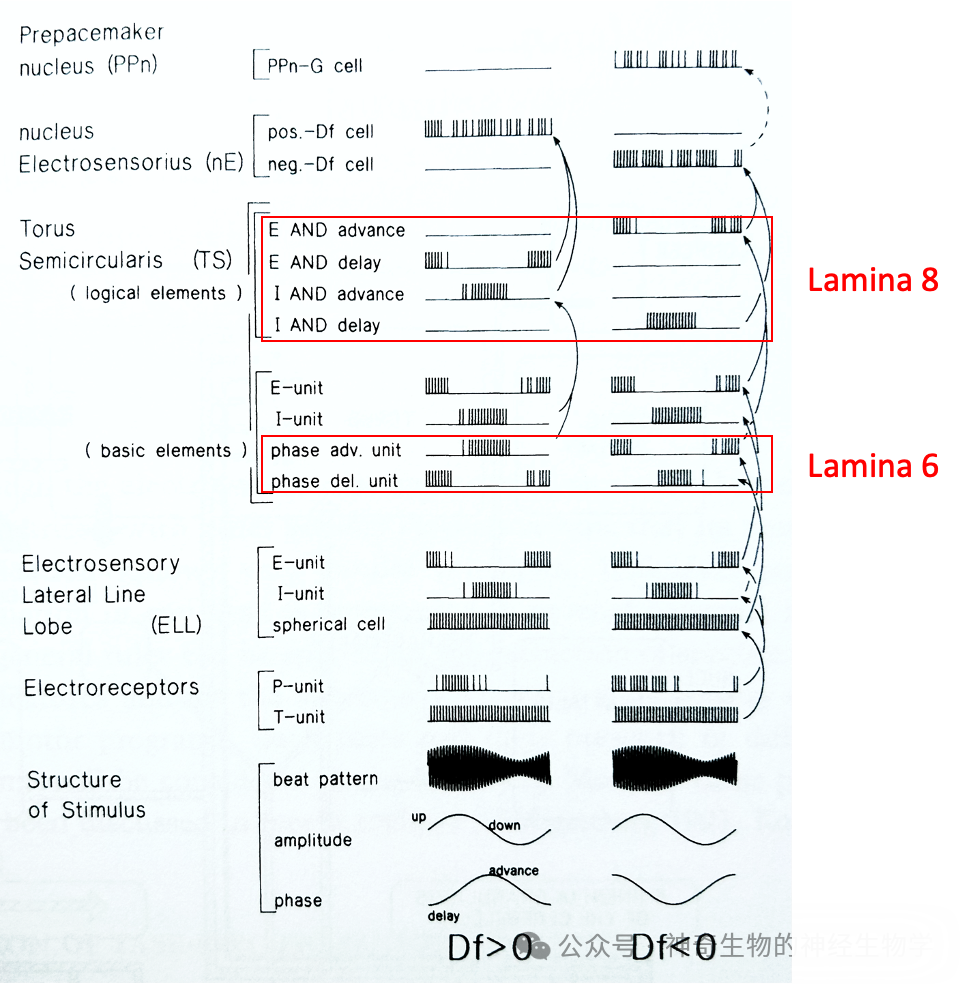
TS输出投射到间脑(diencephalon,即丘脑)核团nE(nucleus electrosensorius),分成两个部分。
(1)nE-up区:背侧(dorsal), TS-2类细胞和TS-3细胞联合激活,得出对方的频率比我慢的结论。注射 L-Glu到这里使得鱼的放电频率缓慢上升,双侧损毁动物失去JAR中上调放电频率的能力。
(2)nE-down:腹侧(ventral), TS-1类细胞和TS-4细胞联合激活,得出对方的频率比我快的结论。注射 L-Glu到这里使得鱼的放电频率缓慢下降,双侧损毁动物失去JAR中下调放电频率的能力。
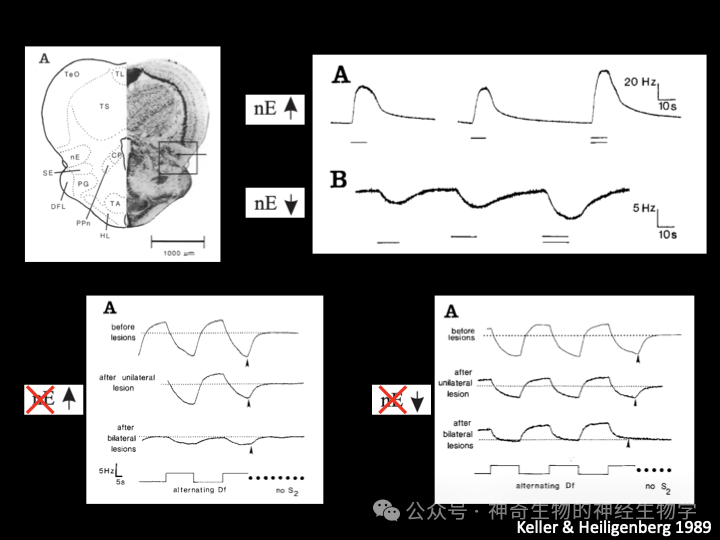
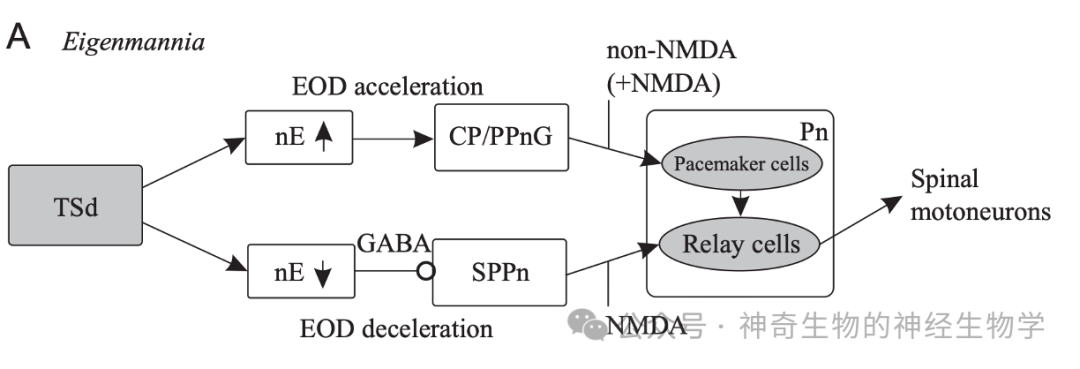
nE下游有前起搏器核团PPn(prepacemaker nucleus)被nE-up激活;提高pacemaker频率,(AMPA-R)。SPPn(sublemniscal prepacemaker nucleus)持续激活后脑的relay cells,抑制或损毁它会降低放电频率,依赖NMDA-R;被nE-down抑制(GABA)。最后一级是后脑(延髓)起搏器pacemaker cell - relay cells:电偶联,直接控制放电器官每一次放电。
交流信号中的时间编码
电鱼除了通过放电来感知环境之外,也可以通过放电来与同类交流。
脉冲型非洲电鱼中,EL(exterolateral nucleus)核团(即正弦波型电鱼中的TS脑区,torus semicircularis,即四足动物脑的下丘inferior colliculus)复杂处理通讯信号里的时间信息。有些物种的EL有两个瓣,ELa和ELp。ELa和ELp分别处理两种尺度的时间信息,通过回放(playback)实验检测这两种信息的重要性。
不同物种的电鱼的电脉冲的波形各不相同。同一个物种的不同个体的电脉冲的波形也略有不同。所以电脉冲的波形是身份的标识,电鱼可以通过它来认出对方是谁。

不同物种的波形差异较大,同一物种的不同性别,不同个体的差异就比较小,微秒(us)尺度的差异。有些物种中,EOD波形长度和个体长度相关。只有clade-A的电鱼,也就是有ELa和ELp分化的物种能区分不同的EOD波形。
EOD波形如何识别呢?假设有一个简单的方波:身体两侧的电感受器,会分别被方波的上升相和下降相激活。身体两侧的电感受器激活的时间差就编码了方波的持续时长。如果是个多相EOD,那么身体各处的电感受器会以不同的latency被EOD不同相激活。
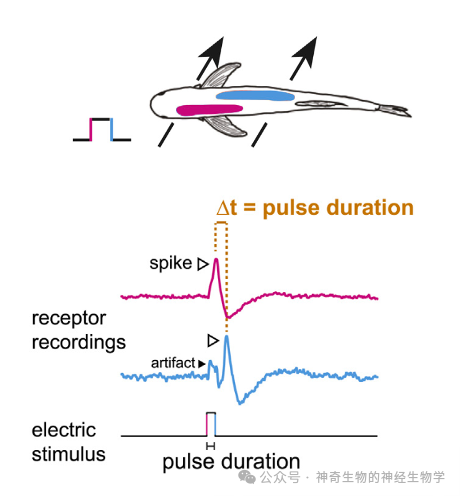
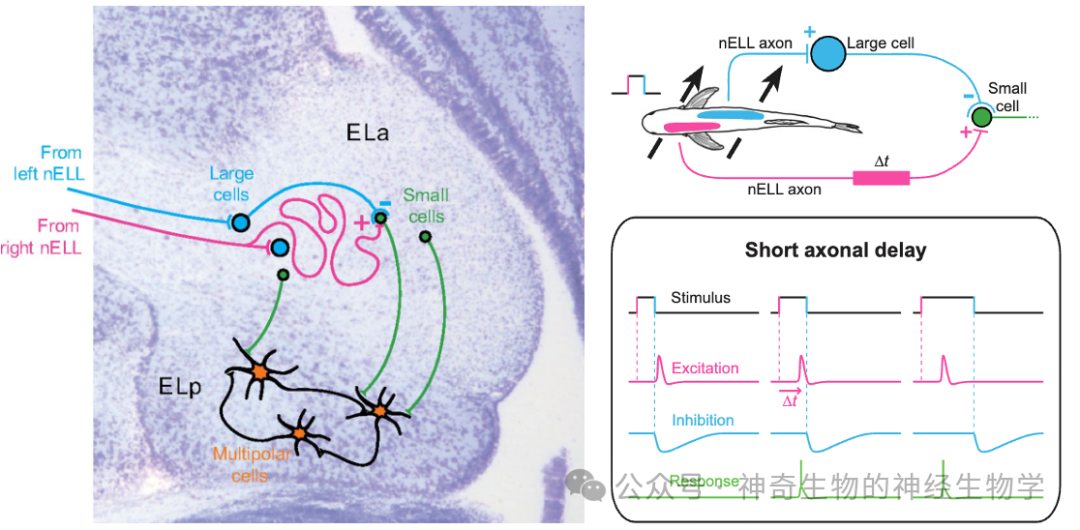
ELa也具备一个延迟线-重合检测器(delay line and coincidence detector):
长延时的激活信号:同侧nELL通过迂回axon激活small cells(检测us尺度的差异。axon传播速度15m/s)。迂回axon沿线激活很多不同的small cells,于是不同的small cells获得不同的传播延时。
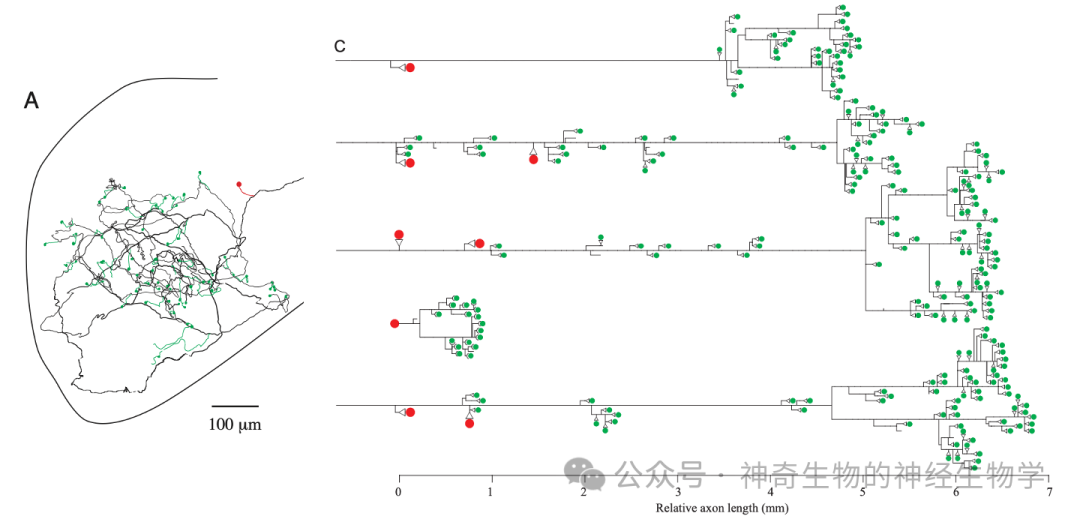
无延时的抑制信号:对侧nELL短axon激活large cells,large cells(抑制性的)通过短axon抑制small cells;抑制的时间常数比较长。
small cell作为重合检测器。只有当左右两侧激活时间差(即EOD的时长)大于axon的延迟,同侧的有延迟激活会早于对侧无延时的抑制信号到到small cell,该small cell才会被激活。迂回axon沿线不同的small cells检测到重合,就能编码出不同的EOD的时长。
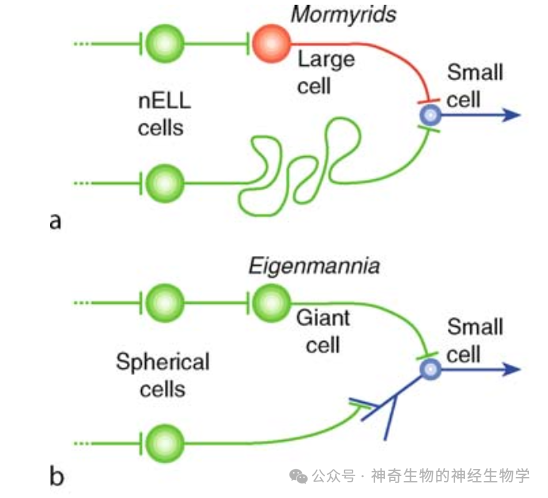
这样的结构正好和正弦波型美洲电鱼在同一脑区(TS lamina 6)的相位比较器是反的。
而电波通过说话的内容则是通过“嗒嗒,嗒嗒,嗒嗒嗒嗒……”的频率变化的表达。虽然我们还不能完全解读电鱼们之间“嗒嗒,嗒嗒嗒嗒……”地都在说些什么,但我们是可以看出他们之间是有交流的,一只电鱼快速的地“嗒嗒嗒嗒嗒嗒”一通后,另外一只电鱼往往也会“嗒嗒嗒嗒嗒嗒” 一通作为回应。就像是摩斯电码。
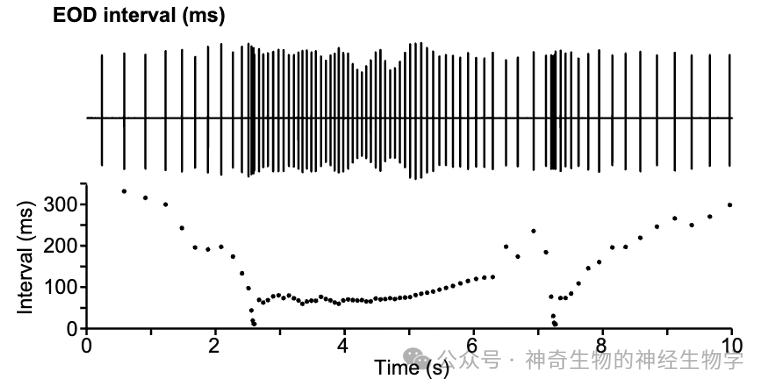
脉冲频率 (或者用interpulse interval,IPI 描述),传递交流的信息。这是在ELp进行处理:
突触通过时间整合、短时可塑性(易化或者衰减)可以呈现出不同的滤波特性,使得一级可以产生对不同脉冲频率的响应偏好(tunning),从而构造出基础的高通滤波器和低通滤波器。
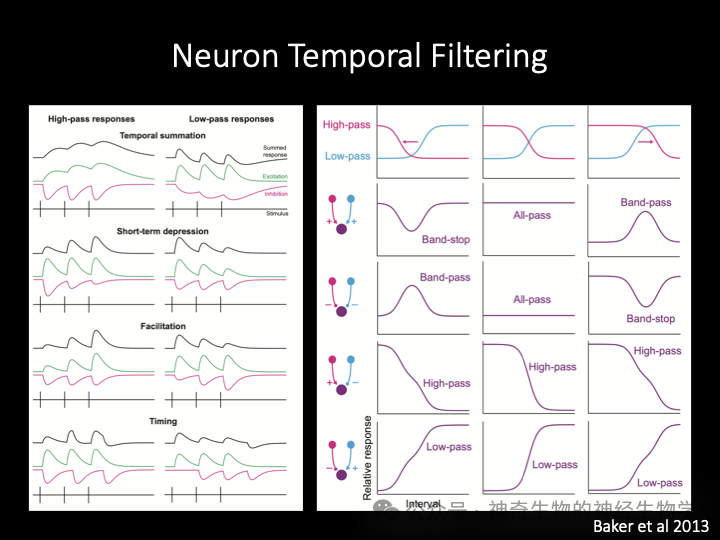
通过兴奋和抑制突触的组合,可以产生更加复杂的组合滤波器和带通滤波器。这也是神经系统对时间序列信息处理的基本原理。
感知副本
前面我们说到,放电来感知环境的时候,电鱼要想办法排除来自同类产生的电波的干扰。而如果是放电来和同类交流,就反过来要排除自己放出电波对接听到别人发来的电波所产生的干扰。
这其实一个神经系统非常普遍的功能,叫做感知副本(efference copy,又称corollary discharge)。我们在说话的时候还能同时听到别人说话的声音,就是因为我们的大脑发出要说什么话的命令给咽喉的同时,也把要说的内容抄送了一份给分析处理听觉的脑区,这样听觉脑区就可以知道此时那些话是我们自己说的,那些话是听到别人说的。
感知副本其实渗透在我们大脑功能的方方面面,比如说我们会在过山车的感觉到天旋地转,而如果是自己抬头看天低头看地左顾右盼,则不会觉得世界被颠倒。这就是因为当我们自己主动转头的时候,我们大脑也会发送一份转头之后看到的世界会怎样旋转的感知副本给我们的视觉脑区。而如果是猝不及防被动转头的话,视觉系统没有事先收到通知,就会真的以为世界颠倒了。
感知副本在我们不经意间发生,但仔细想想其实是很复杂一种操作。大脑要如何生成那个感知副本呢?转头的命令本身是控制脖子肌肉如何收缩,大脑如何把肌肉收缩转写成眼睛看到图像的旋转?柯蒂斯·贝尔(Curtis Bell)在1980年代做了一个精妙绝伦的实验,在神经算法层面上第一次完整地解释电鱼在交流的过程中的感知副本是如何生成的。
贝尔的研究是在发送电脉冲的非洲电鱼里做的。他首先麻痹了电鱼的放电器官,使它无法放电而不自知。然后用一根电极记录大脑向放电器官发出的放电命令,每当大脑发出一个放电命令,贝尔就在水里用刺激电极生成一个人工的波形(方波),这样就移花接木地把电鱼自己产生的电脉冲替换成了一个人工的方波,与此同时记录电鱼大脑中感受电信号的核团(ELL)的反应。
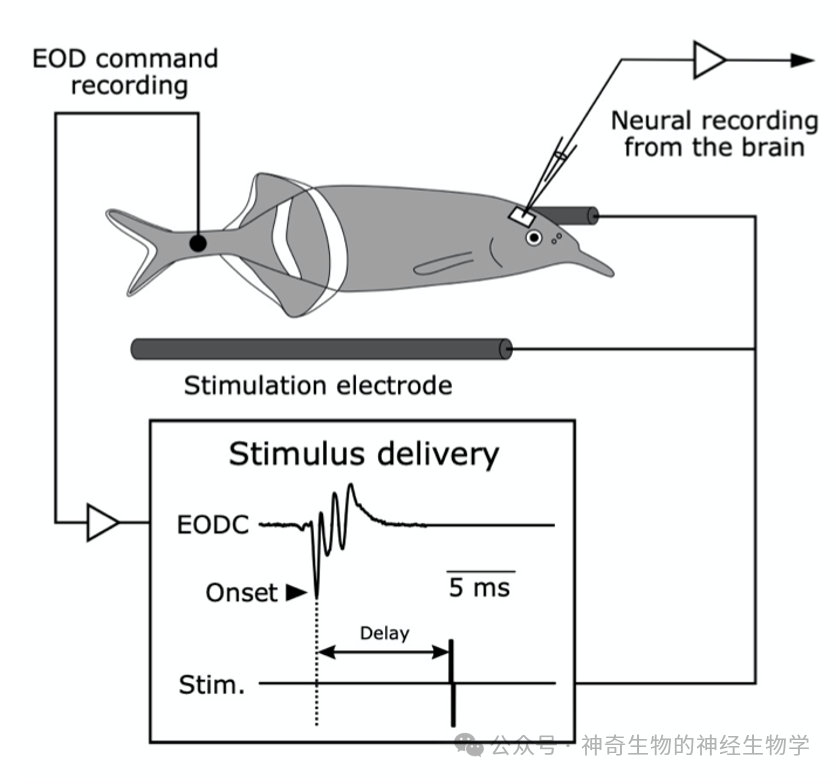
贝尔发现,一开始电鱼对方波的反应就是一个正常的方波。但渐渐地,不到10分钟之后,这个反应就消失了!此时如果停止播放方波,每次电鱼的大脑发送一次放电命令(因为放电器官被麻痹而实际没有放电),电感觉核团就会出现一个反向的方波!
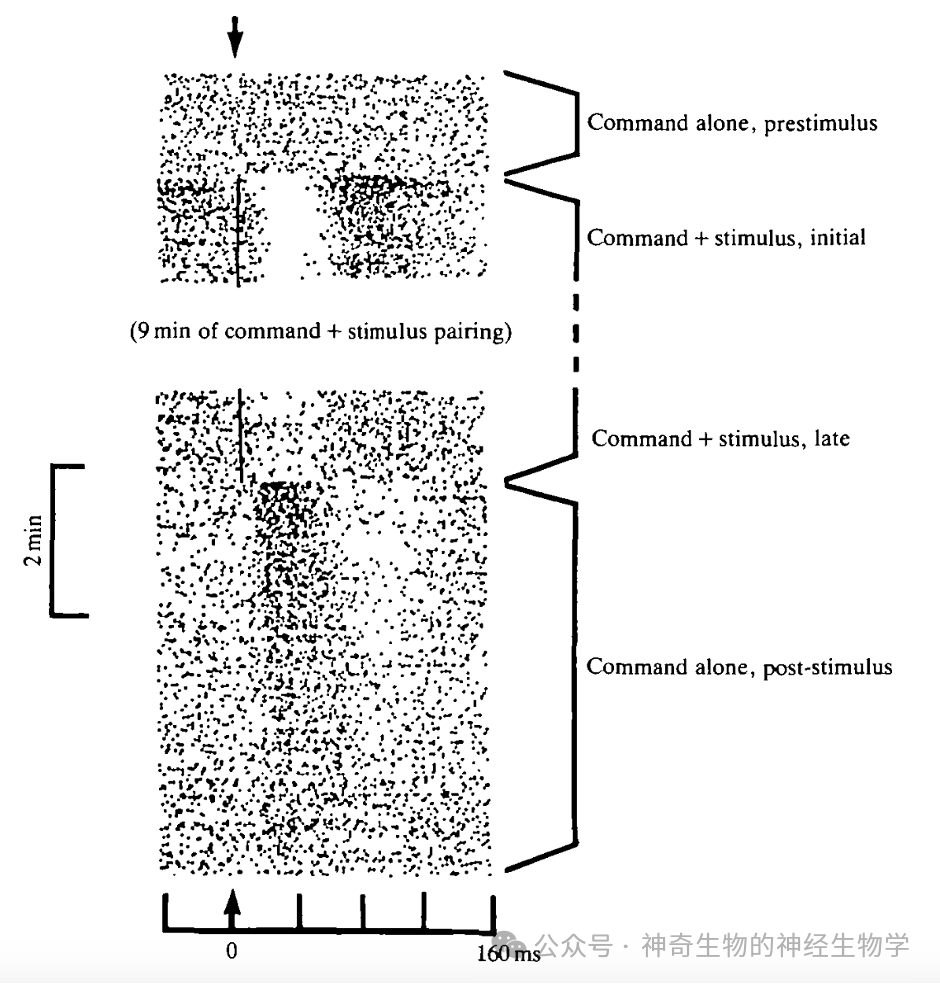
也就是说,之前对于方波反应的消失是因为,大脑每次发送放电命令的时候,同时发送了一份感知副本给电感觉核团,而这个感知副本就是一个反向的方波,所以可以抵消到人工生成的那个正向的方波!电鱼在短时间内就接受了自己产生的电脉冲变成了方波这样安排,然后主动地更新了自己的感知副本。
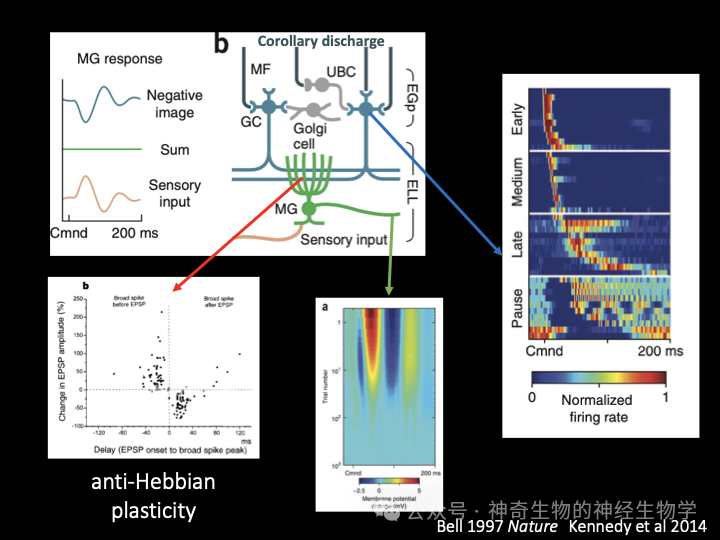
贝尔进一步的研究发现,电鱼的后脑的电感觉核团有十分类似于小脑的结构:大量颗粒细胞形成平行纤维,和少量的浦肯野细胞建立连接。来自**区域的放电命令激活颗粒细胞,每个颗粒细胞对放电命令进行不同的滤波和变形,大量颗粒细胞可以产生出各种各样的波形,浦肯野细胞通过比较哪个波形的排列组合最能抵消掉感觉细胞接受的波形,最后反向强化(anti-Hebbian plasticity)了与那些能够产生电脉冲波形的颗粒细胞的连接。通过这种办法,电鱼就可以动态的生成反向波形来抵消自己产生的电脉冲在脑中的反应。这就像是主动降噪耳机的工作原理,只是耳机收集的是环境音,然后持续生成反向的波形与之相抵,而电鱼收集的是自己的声音,而只在自己说话的时候开启降噪功能。
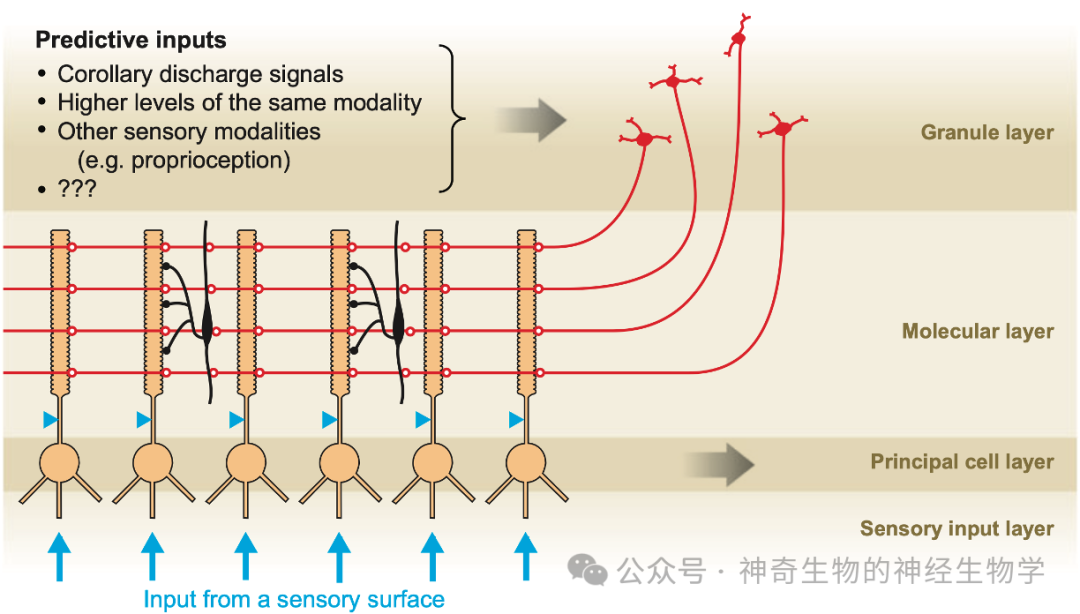
这就是感知副本的生成原理,也广泛适用于其他动物的其他模态的感觉**整合。这样的神经网络的学习过程,可以拟合任意曲线,可以学习任意**指令可能造成的感觉输入的变化的关系。神经系统很难建立起一个外部世界的物理模型,根据力学定律预测**输出会造成什么样的感觉变化:比如喉部肌肉的收缩指令和发出声音的声学特征之间关系不是通过大脑内部的物理模型来解算,而是基于无数次训练学到的统计规律(其实和AI是一样的)。
参考文献
Heiligenberg, W. (1991). Neural nets in electric fish. MIT press.
Watanabe, A., & Takeda, K. (1963). The change of discharge frequency by AC stimulus in a weak electric fish. Journal of Experimental Biology, 40(1), 57-66.
Bullock, T. H., Hamstra, R. H., & Scheich, H. (1972). The jamming avoidance response of high frequency electric fish: I. General features. Journal of comparative physiology, 77, 1-22.
Partridge, B. L., Heiligenberg, W., & Matsubara, J. (1981). The neural basis of a sensory filter in the jamming avoidance response: no grandmother cells in sight. Journal of comparative physiology, 145(2), 153-168.
Heiligenberg, W., & Rose, G. (1985). Phase and amplitude computations in the midbrain of an electric fish: intracellular studies of neurons participating in the jamming avoidance response of Eigenmannia. Journal of Neuroscience, 5(2), 515-531.
Carr, C. E., Heiligenberg, W., & Rose, G. J. (1986). A time-comparison circuit in the electric fish midbrain. I. Behavior and physiology. Journal of Neuroscience, 6(1), 107-119.
Carr, C. E., Maler, L., & Taylor, B. (1986). A time-comparison circuit in the electric fish midbrain. II. Functional morphology. Journal of Neuroscience, 6(5), 1372-1383.
Heiligenberg, W., Baker, C., & Matsubara, J. (1978). The jamming avoidance response in Eigenmannia revisited: the structure of a neuronal democracy. Journal of comparative physiology, 127, 267-286.
Rose, G. J. (2004). Insights into neural mechanisms and evolution of behaviour from electric fish. Nature Reviews Neuroscience, 5(12), 943-951
Baker, C. A., Kohashi, T., Lyons-Warren, A. M., Ma, X., & Carlson, B. A. (2013). Multiplexed temporal coding of electric communication signals in mormyrid fishes. Journal of Experimental Biology, 216(13), 2365-2379.
Xu-Friedman, Matthew A., and Carl D. Hopkins. "Central mechanisms of temporal analysis in the knollenorgan pathway of mormyrid electric fish." Journal of Experimental Biology 202.10 (1999): 1311-1318.
Friedman, M. A., & Hopkins, C. D. (1998). Neural substrates for species recognition in the time-coding electrosensory pathway of mormyrid electric fish. Journal of Neuroscience, 18(3), 1171-1185.
Bell, C. C. (1981). An efference copy which is modified by reafferent input. Science, 214(4519), 450-453.
Bell, C. C. (1989). Sensory coding and corollary discharge effects in mormyrid electric fish. Journal of Experimental Biology, 146(1), 229-253.
Bell, C. C., Han, V. Z., Sugawara, Y., & Grant, K. (1997). Synaptic plasticity in a cerebellum-like structure depends on temporal order. Nature, 387(6630), 278-281.
Bell, C. C., Han, V., & Sawtell, N. B. (2008). Cerebellum-like structures and their implications for cerebellar function. Annu. Rev. Neurosci., 31, 1-24.

 美国
美国 加拿大
加拿大 新加坡
新加坡 日本
日本 迪拜
迪拜 澳洲
澳洲 泰国
泰国 越南
越南 新西兰
新西兰 马来
马来 意大利
意大利 英国
英国 德国
德国 法国
法国 西班牙
西班牙





